Article Text

Abstract
Asthma is a very heterozygous disease, divided in subtypes, such as eosinophilic and neutrophilic asthma. Phenotyping and endotyping of patients, especially patients with severe asthma who are refractory to standard treatment, are crucial in asthma management and are based on a combination of clinical and biological features. Nevertheless, the quest remains to find better biomarkers that distinguish asthma subtypes in a more clear and objective manner and to find new therapeutic targets to treat people with therapy-resistant asthma. In the past, research to identify asthma subtypes mainly focused on expression profiles of protein-coding genes. However, advances in RNA-sequencing technologies and the discovery of non-coding RNAs as important post-transcriptional regulators have provided an entire new field of research opportunities in asthma. This review focusses on long non-coding RNAs (lncRNAs) in asthma; these are non-coding RNAs with a length of more than 200 nucleotides. Many lncRNAs are differentially expressed in asthma, and several have been associated with asthma severity or inflammatory phenotype. Moreover, in vivo and in vitro functional studies have identified the mechanisms of action of specific lncRNAs. Although lncRNAs remain not widely studied in asthma, the current studies show the potential of lncRNAs as biomarkers and therapeutic targets as well as the need for further research.
- asthma
- asthma genetics
- asthma mechanisms
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
Almost 340 million people worldwide suffer from asthma. Cases are found with a higher frequency in Western and more industrialised countries, where up to 10% of the population has asthma.1 Asthma is not one well-defined disease. Asthma symptoms include wheezing, chest tightness, coughing, shortness of breath and airflow limitation. These symptoms can change over time and do not affect all patients with similar intensity or frequency. This is due to both genetic and environmental factors which influence the pathogenic alterations within the airways, such as inflammation, airway remodelling, mucous hypersecretion, airway hyper-responsiveness and reversible airway obstruction. To describe the heterogeneity of asthma, several subtypes are defined. While multiple classification systems exist, the subtypes defined by Wenzel were mostly used in the studies discussed further.2 According to Wenzel, there are six different forms of asthma : (1) early-onset allergic Th2, (2) late-onset eosinophilic, (3) exercise-induced, (4) obesity-related, (5) neutrophilic and (6) paucigranulocytic asthma. Cases are sorted into the neutrophilic, eosinophilic and paucigranulocytic groups according to the percentage of inflammatory cells present in the airways. Although neutrophils represent the largest fraction of inflammatory cells in the airways, patients are categorised as having eosinophilic asthma if their sputum contains more than 3% eosinophils. Neutrophilic asthma is defined by more than 61% neutrophils in sputum and paucigranulocytic asthma by less than 3% eosinophils and less than 61% neutrophils in sputum.3 Still, these cut-offs—especially those for neutrophils—can differ among different research groups and geographical locations. The different asthma categories have now been further defined based on the underlying biology thanks to the advances in molecular techniques. Nevertheless, treatment response can overlap and a need for biomarkers to distinguish these subtypes in a more objective and practical manner remains.2
The immunological mechanisms in asthma are best understood for early-onset allergic asthma, where allergen exposure in susceptible individuals induces the initiation of a Th2 response, in which type 2 cytokines interleukin (IL)-4, IL-13 and IL-5 induce IgE production and activation of mast cells, recruitment of eosinophils, formation of mucus-producing goblet cells and thickening of the airway smooth muscle cells (ASMCs).4 5 These type 2 cytokines can also be produced by innate lymphoid cells type 2, which are considered to be important in late onset-eosinophilic asthma.5 In neutrophilic asthma, other pathways are thought to be of importance, including the induction of Th1 and Th17 responses.2 4
Standard-of-care treatment for asthma consists of inhaled corticosteroids (ICS). Administration of corticosteroids results in the suppression of proinflammatory gene expression, inactivation of Th2 cells, mast cells and eosinophils, and an increase of anti-inflammatory gene expression as well as activation of regulatory T cells (Tregs).6 7 However, ICS are less efficient to control severe asthma and—when taken chronically—can lead to serious side effects, such as osteoporosis, diabetes and reduced growth velocity.8 Other treatments include long-acting inhaled β2 agonists, low-dose theophylline, antileukotrienes and—for severe asthma—also biologicals. While all of these agents can bring substantial relief to patients, none of them are curative, and some patients appear to be insensitive to these and similar medicines.7 9 Despite the successes of type 2-targeting biologicals to treat severe eosinophilic asthma, there remains an unmet need for additional therapeutic options covering the range of patient-specific requirements, particularly for severe ICS-resistant patients with asthma. A crucial step to achieve such precision medicine is the discovery of accurate biomarkers to distinguish different disease phenotypes and to determine optimal therapy (predictive biomarker) or predict the risk of future exacerbations (prognostic biomarker). Thus far, most biomarker studies have focused on protein-coding genes. However, the potential of non-coding RNAs as new biomarkers remains underexplored.10–15
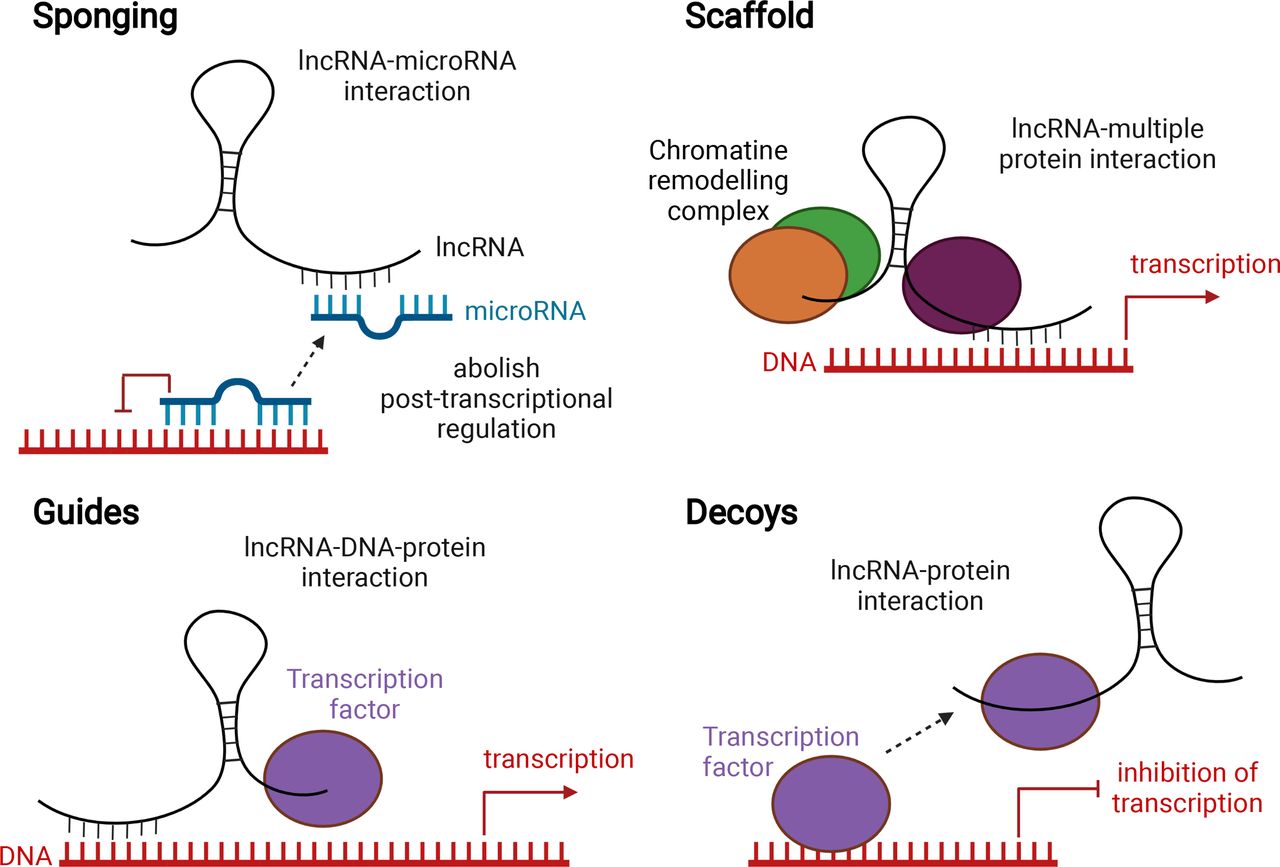
The advances of RNA-sequencing technology revealed that our genome is pervasively transcribed producing thousands of coding and non-coding RNA molecules (ncRNAs). ncRNAs are transcribed, but not translated into proteins.16 There are different subtypes of ncRNAs, including microRNAs, ribosomal RNAs and transfer RNAs. In this review, we will focus on long non-coding RNAs (lncRNAs), as the role of microRNAs in asthma has been extensively studied and reviewed.17–20 lncRNAs are arbitrarily defined by their length, over 200 nucleotides, and are emerging as regulators of various cellular processes, making them possible biomarkers and therapeutic targets.21 The origin of lncRNAs is not always known. Some are conserved among species; others are derived from protein-coding genes as a result of frame disruptions, chromosome rearrangements, duplication of another non-coding gene by retrotransposition, tandem duplication or insertion of a transposable element.22 Although lncRNAs generally have a low sequence conservation, their structure, splicing pattern and subcellular localisation are often conserved.23 lncRNAs can function through a variety of mechanisms and are defined into several functional subtypes, among which the sponges, scaffolds, guides and decoys are among the most reported ones (figure 1)24–27. Sponging lncRNAs can bind to microRNAs, thus preventing these microRNAs, which control post-transcriptional processes, to interact with their targets. lncRNAs can also facilitate the formation of chromatin remodelling complexes and subsequent gene transcription (scaffold). lncRNAs can function as guides to lead a transcription factor to a specific DNA location to regulate transcription. As decoys, lncRNAs can bind a transcription factor, preventing it from binding to DNA (figure 1). The potential involvement of lncRNAs in the pathogenesis of several diseases has been reported.28–31

Main proposed working mechanisms of lncRNAs. lncRNAs can act as a sponge for microRNAs. By binding to these microRNAs, they prevent microRNAs from binding to their targets, thereby abolishing post-transcriptional regulation (top left). lncRNAs can act as scaffolds to facilitate the assembly of chromatin remodelling complexes and subsequent transcription (top right). lncRNAs can guide transcription factors to specific locations on the genome, thus facilitating transcription (bottom left). lncRNAs can act as a decoy by binding to transcription factors, thus leading to transcriptional repression (bottom right). Created with BioRender.com. lncRNA, long non-coding RNA.
This review discusses the status of lncRNA research in asthma and defines the next steps required to evaluate their potential as new biomarkers or therapeutic targets. PubMed was searched using the query “lncRNAs” AND “asthma”. Articles regarding lncRNA discovery in asthma as well as in vitro and in vivo functional studies published since 2012 were included.
Identifying lncRNAs in asthma
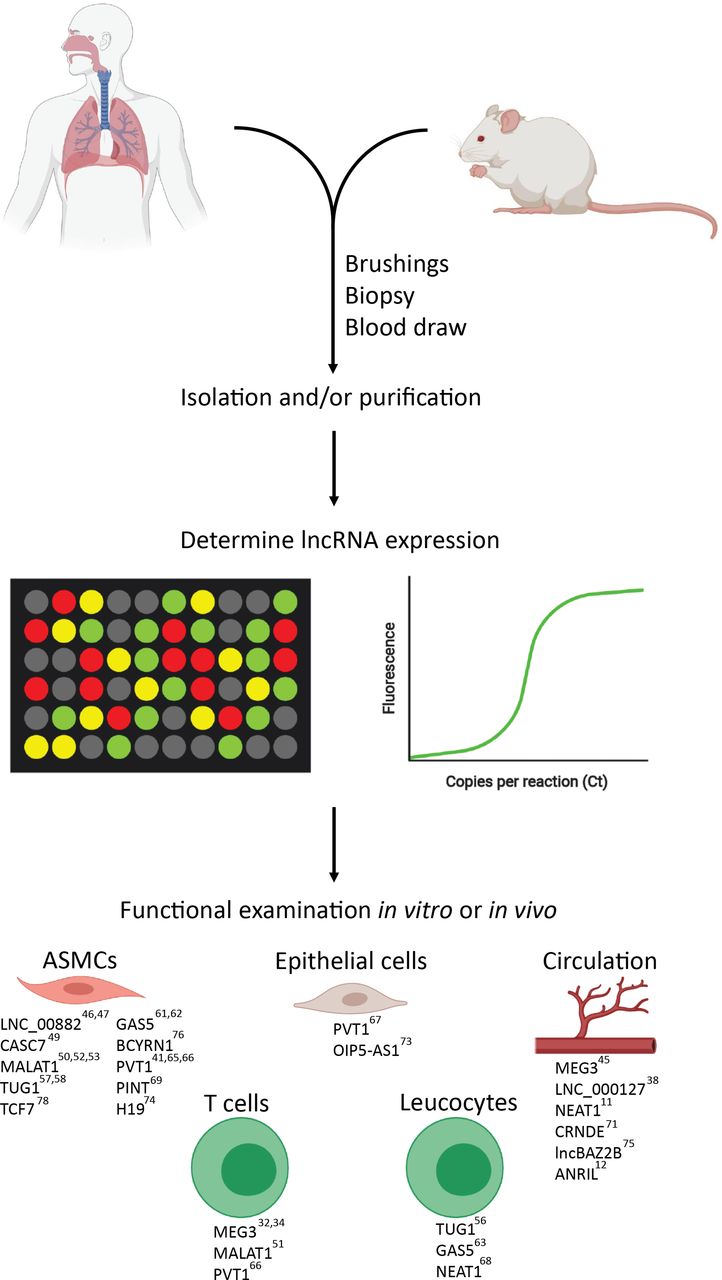
To identify asthma-associated lncRNAs, gene expression profiling technologies, such as RNA-sequencing or microarrays, are typically used. There are multiple relevant cell types in asthma, such as immune cells and respiratory epithelial cells, which can be either sampled in blood, sputum or by bronchial brushings. The following paragraphs discuss human studies where lncRNA expression in samples from patients with asthma and healthy individuals were compared. Significant differential lncRNAs are typically selected based on a statistical test and resulting p value or false discovery rate (FDR). Also the fold change (FC), which shows the extent of differential expression, is often used. Sample size and applied cut-off values (both for p value, FDR and FC) will affect the number of identified differentially expressed lncRNAs. Since many discovery studies are underpowered, with small sample sizes (table 1), we focused on those that found lncRNAs with functions that can contribute to asthma pathogenesis. The differences in technical approaches and cut-off values, sample type and size and asthma heterogeneity are important to keep in mind when comparing results between studies.
Summary of human studies with gene expression profile data in asthma focusing on lncRNAs
lncRNAs in T cells
CD4+ T cells drive the inflammatory and allergic responses in allergic asthma. Three independent studies have profiled lncRNA expression in CD4+ T cells in human peripheral blood T cells.32–34 Notably, the number of differentially expressed lncRNAs between CD4+ T cells from asthma samples and healthy controls differs strongly between different studies, ranging from 6 to over 2000 (table 1). Although differential lncRNA expression is confirmed by reverse transcription quantitative PCR (RT-PCR) within specific studies, there is unfortunately no overlap in the reported lncRNAs from the different studies, which may be due to differences in asthma phenotype, source of the T cells, sample number and used technologies,32–35 as well as the difficulty to evaluate overlap due to annotation discrepancies.
Still, some studies linked the identified lncRNAs to asthma severity or pathogenesis. Tsitsiou et al, for example, did find not only 6 differentially expressed lncRNAs in CD4+ T cells but also 167 differentially expressed lncRNAs in CD8+ T cells in patients with severe asthma (n=12) compared with healthy controls (n=8).32 Moreover, these CD8+ T cells, but not CD4+ T cells, were activated in severe asthma, suggesting an involvement of lncRNAs in CD8+ T-cell activation in severe asthma.32 This novel concept requires further validation.
In a large study, Qiu et al identified 225 differentially expressed lncRNAs in CD4+ T cells from severe asthma (n=52) compared with controls (n=45),34 and demonstrated that the lncRNA MEG3 plays a role in Treg/Th17 balance, which has been linked to allergic asthma before.36
The functional role of MEG3 and other identified lncRNAs (eg, MALAT-1 and CACS7)34 will be further discussed in the next section.
lncRNAs in blood
Other studies focus on lncRNA expression in whole blood rather than individual cell types isolated from blood.37–39 Whole blood can be of interest to look for biomarkers, as there can be spillover of inflammatory processes from the lung into the blood, and it is easy to implement the sampling in the clinic. Nevertheless, whole blood also contains lncRNAs associated with the blood immunological profile as such, being not-related to asthma, as well as platelet-derived lncRNAs.
Zhu and colleagues investigated the lncRNA expression profiles in eosinophilic asthma.39 They found 41 differentially expressed lncRNAs in whole blood from eosinophilic asthma (n=9) compared with controls (n=3). Pathway analysis based on the coexpression of certain lncRNAs with certain messenger RNA (mRNAs) revealed that the 41 lncRNAs were mainly correlated to mRNAs involved in immune regulation.39 Moreover, these transcripts could be detected in peripheral blood, supporting their role as potential biomarkers, yet further work will be needed in larger and independent cohorts to confirm their diagnostic/prognostic value in eosinophilic asthma. The same research group subsequently investigated whether lncRNA expression differs between asthma inflammatory phenotypes and found 190 differentially expressed lncRNAs in eosinophilic asthma (n=12) and 166 in neutrophilic asthma (n=6) compared with healthy controls (n=6). Notably, LNC_000127 was expressed higher in eosinophilic asthma compared with neutrophilic asthma and controls (more details are discussed in the online supplemental materials).38
lncRNAs in airways
Finally, the lncRNA expression landscape was also investigated in samples from the airways. Liu et al looked at publicly available datasets (NCBI Gene Expression Omnibus and GEO) from airway epithelial brushings from patients with asthma (n=62) and controls (n=43) and identified 159 differentially expressed lncRNAs between these two groups.40 The authors then selected eight lncRNAs that potentially interact with mRNAs, based on functional gene analysis through coexpression networks of lncRNAs and mRNAs. These eight lncRNAs were enriched in asthma-associated pathways such as airway remodelling and vascular endothelial growth. Experimental studies are required to validate interactions between these lncRNAs and mRNAs.
Austin et al used microarrays to look at differential lncRNA expression in ASMCs. Compared with healthy controls (n=9), 21 lncRNAs were differentially expressed in non-severe asthma (n=9) and 19 lncRNAs were differentially expressed in severe asthma (n=9). Four of the lncRNAs were overlapping, including PVT1, which will be further discussed.41
Boudewijn et al performed RNA-sequencing on bronchial biopsies of patients with asthma who were in remission (n=14) and patients with asthma with persistent asthma (n=46). They found 35 lncRNAs that were linked with complete remission, yet no information on expression patterns was provided.42 Understanding the role of these lncRNAs may provide important insights into the disease trajectory of asthma.
In summary, the aforementioned studies all identified multiple lncRNAs exhibiting differential expression in asthma or specific asthma phenotypes, but few lncRNAs were replicated in different studies (table 1). This can probably be explained by the small sample cohorts in these studies and the huge difference in the origin of biological samples.
Functional role of lncRNAs in asthma
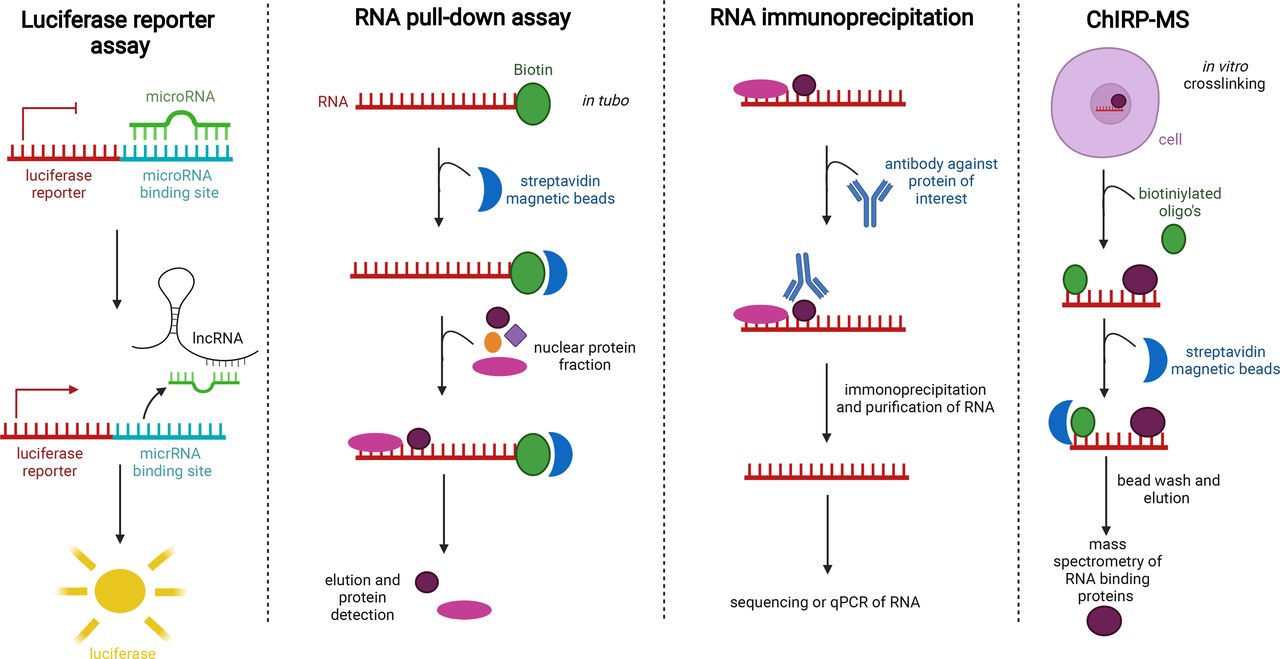
The functional role of several specific lncRNAs, which were selected based on profiling studies (see above) or on previous knowledge in other diseases, was more thoroughly investigated in several studies. In the following paragraphs, we focus on specific lncRNAs that are either upregulated or downregulated in asthma and were functionally studied. We describe the in vitro and/or in vivo approaches that were used to obtain insights in lncRNA function. An overview of these lncRNAs, their expression in asthma, their interactions with miRNAs or proteins and their potential role in specific asthma-associated processes is given in table 2 and figure 2. Techniques to study interactions between lncRNAs and proteins or RNA are shown in figure 3. These include luciferase reporter assays to visualise sponging of microRNAs and RNA pull-down experiments to detect proteins and RNAs that were bound to an RNA of interest. Also RNA immunoprecipitation helps to detect RNAs that bind to a protein of interest.
lncRNAs and their possible interaction partners and effects, based on human and animal data

lncRNAs differenally expressed in asthma in human, rat or mice in different cell types. Created with BioRender.com. ASMC, airway smooth muscle cell; lncRNA, long non-coding RNA.

{kind=link}
{kind=link}
{kind=link}
Different techniques to study protein–RNA and RNA–RNA interactions. With a luciferase reporter assay, sponging of a microRNA can be visualised. RNA pull-down enables us to extract proteins that bind to an RNA sequence of interest. RNA immunoprecipitation makes it possible to detect RNA that binds to a certain protein of interest. ChIRP-MS enables us to perform mass spectrometry on proteins that bind to an RNA sequence of interest. lncRNA, long non-coding RNA.
Maternally expressed 3 (MEG3)
Although MEG3 was identified as differentially expressed in several studies, the data are not univocal. In CD4+ T cells of patients with severe asthma, MEG3 expression levels were upregulated and associated with reduced microRNA-17 expression.34 Silencing of MEG3 expression in CD4+ T cells by siRNA knockdown reduced the expression of Th17 transcription factor RORγt and IL-17 and resulted in upregulation of microRNA-17. Luciferase reporter assays and RNA pull-down experiments (figure 3) demonstrated that MEG3 inhibits microRNA-17 by functioning as a competing RNA sponge. Moreover, microRNA-17 can suppress Th17 response by targeting RORyt, suggesting that MEG3 plays a role in the Th17 imbalance in asthma.34 Since elevated Th17 responses are linked to moderate and severe asthma, MEG3 could be a biomarker for asthma severity.43
In CD8+ T cells, on the other hand, MEG3 was found downregulated.32 This contradictory finding requires further research and validation, since RORyt+ CD8+Tc17 cells that produce IL-17 have been proposed to support Th17 responses in asthma.44 MEG3 was also downregulated in serum of patients with asthma compared with controls, especially in mixed eosinophilic/neutrophilic asthma, and was negatively correlated with asthma progression.45 Possibly the different sample types and timing can influence the findings on MEG3 expression in asthma.34 45
LNC_00882
lncRNAs can also affect airway smooth muscle function, which is important in asthma.46 The role of LNC_00882 was evaluated through gain-of-function and loss-of-function experiments in ASMCs. LNC_00882 was elevated in ASMCs treated with platelet-derived growth factor (PDGF) and promoted their proliferation. Luciferase reporter and RNA pull-down assays to discover the interaction partners of LNC_00882 revealed that LNC_00882 directly interacted with microRNA-3169-5p that can target β-catenin. Liu et al thus concluded that LNC_00882 promotes the proliferation of ASMCs by enhancing Wnt/β-catenin signalling via sponging of microRNA-3619-5p, supporting a role for LNC_00882 in airway remodelling.46
Perry et al also observed upregulation of LNC_00882 in human ASMCs and that this lncRNA has seven sites that are complementary to microRNA-371.47 Notably, others reported that microRNA-371 (in cooperation with four other microRNAs) plays a role in the Th1/Th2 balance in asthma by inhibiting Runx3, a transcription factor that promotes Th1 diferentiation.48 It remains to be investigated whether LNC_00882 and/or microRNA-371 fulfil similar roles in ASMCs.
Cancer susceptibility candidate 7 (CASC7)
In ASMCs from patients with severe asthma, Liu et al discovered that CASC7 expression was suppressed49 and that it has a binding site for microRNA-21, a miRNA that is part of signalling pathway together with PTEN and AKT. Both microRNA-21 and AKT activity were upregulated in ASMCs from patients with severe asthma. Modulation of CASC7 and microRNA-21 expression and luciferase reporter assays demonstrated interactions between CASC7, microRNA-21 and PTEN mRNA. Overall, Liu et al concluded that CASC7 promotes PTEN expression through inhibiting microRNA-21 expression. Moreover, overexpression of CASC7 inhibited the PI3K/AKT pathway leading to increased corticosteroid sensitivity.49 Further in vivo experiments combining corticosteroids and CASC7 RNA could evaluate therapeutic potential. Of note, in CD4+ T cells, an upregulated expression of CASC7 was reported.34 Since lncRNAs can have cell type-specific functions, these two studies are not necessarily contradictory.
Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1)
Four studies observed higher expression of MALAT1 in asthma in human ASMCs induced by PDGF,50 in CD4+ T cells isolated from blood from patients with asthma,51 in ASMCs isolated from asthmatic rats52 and in newborn asthmatic rat models.53 In human ASMCs, siRNA-mediated knockdown of MALAT1 inhibited proliferation and migration.50 Luciferase reporter assays revealed a direct interaction between MALAT1 and microRNA-150. The translation initiation factor 4E (eIF4E), a key player in the AKT pathway, is a direct target of microRNA-150. As a result, MALAT1 can activate AKT signalling through sponging of microRNA-150, leading to proliferation and migration of ASMCs (airway remodelling).50 Another independent in vitro study in rat ASMCs associated MALAT1 levels and microRNA-216a to cell proliferation, migration and apoptosis, suggesting that the interaction of MALAT1 with several miRNAs has a role in airway remodelling.52 Knocking down MALAT1 in primary bronchial and tracheal smooth muscle cells also reduced apoptosis and proinflammatory cytokine production, with involvement of the microRNA-133a-ryanodine receptor-2 (RyR2) axis.53 Notably, RyR2 has been associated with childhood asthma.54
In human CD4+ T cells, Liang et al demonstrated using knockdown experiments and luciferase reporter assays that MALAT1 can also sponge the asthma associated microRNA-155,55 which affects the Th1/Th2 balance in asthma through a CTLA-4-dependent mechanism.51
Overall, MALAT-1 appears to have multifaceted and cell-type specific functions, with involvement in both airway remodelling and the Th1/Th2 balance, supporting further investigations into MALAT1 as therapeutic target for asthma.50 51
Taurine upregulated 1 (TUG1)
Sun et al observed that TUG1 is differentially expressed in monocytes of children with asthma. By performing in vitro experiments on macrophages, they demonstrated that TUG1 sponges microRNA-29c, which has been linked to Th2 cell differentiation.56
By using a rat model, Lin et al studied the function of TUG1 in asthma.57 TUG1 was upregulated in the ASMCs of an asthma rat model compared with controls. In vitro silencing of TUG1 in these asthmatic ASMCs by siRNA led to impaired proliferation and migration and reduced fibroblast growth factor 1 (FGF1) expression. Further analysis demonstrated that TUG1 sponges microRNA-590-5p, which therefore no longer inhibits its target FGF1, thus favouring proliferation and migration of ASMCs (airway remodelling).57
In a study using asthmatic mice, Huang et al could link TUG1 to microRNA-181b. TUG1 acts as a sponge for microRNA-181b, which normally has a protective function in asthma, as it has anti-inflammatory properties.58 However, by sponging, TUG1 inhibits this function and activates the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signalling pathway and promotes airway remodelling.58 Two recent studies confirmed the involvement of TUG1 in ASMC proliferation in human asthma, by impacting on the miR216a-SMURF2 axis and the miR-138–E2F3 axis.59 60
Growth arrest-specific 5 (GAS5)
Multiple studies have looked at the possible function and potential of GAS5.61–63 Wu et al found increased expression of GAS5 in peripheral blood of patients with severe and non-severe asthma compared with healthy controls.63 GAS5 expression reduced after treatment with dexamethasone, a synthetic glucocorticosteroid, in a mouse model of steroid-insensitive asthma. Keenan et al evaluated if GAS5 functioned as a decoy for the glucocorticoid receptor, playing an important role in glucocorticoid sensitivity. Instead of investigating GAS5 expression patterns in asthma, authors examined glucocorticoid sensitivity in vitro and observed that GAS5 can repress glucocorticoid receptor activity of ASMCs.62 Combining the data of Wu et al and Keenan et al, we found that GAS5 seems to promote glucocorticoid insensitivity.62 63
Zhang et al demonstrated that GAS5 expression levels in ASMCs from asthmatic rats are higher than in controls.61 Through RNA immunoprecipitation and RNA pull-down (figure 3), it was demonstrated that GAS5 acts as a sponge for microRNA-10a and that luciferase reporter assays subsequently showed an interaction between microRNA-10a and BDNF. This BNDF protein is produced by ASMCs and enhances inflammation and airway contractility.64 Elevated GAS5 expression promoted ASMC proliferation in vitro through the microRNA-10a/BDNF regulatory axis. Knockdown of GAS5 using siRNAs in vivo decreased airway hyper-responsiveness in asthmatic rats, indicating therapeutic potential.61
Plasmacytoma variant translocation 1 (PVT1)
Comparison of the lncRNA profile in ASMCs from healthy subjects, non-severe corticosteroid sensitive asthma and severe corticosteroid-insensitive asthma identified a number of differentially expressed lncRNAs, including PVT1, which is upregulated in corticosteroid-insensitive asthma.41 After in vitro treatment of these ASMCs with fetal calf serum (FCS) and dexamethasone, Austin et al saw that PVT1 expression was different in the ASMCs originating from the different groups. Whereas PVT1 expression reduced in ASMCs from non-severe asthma, it increased in ASMCs from corticosteroid-insensitive severe asthma and was unaffected in ASMCs from healthy subjects.41 Using siRNA targeting PVT1 in primary smooth muscle cells from patients with severe asthma, Austin et al further observed a decrease of IL-6 release and an increase in ASMC proliferation (airway remodelling).41
Yu et al used a rat model with respiratory syncytial virus (RSV) induced asthma and found PVT1 expression was higher in ASMCs.65 Treatment with α-asarone, an anti-inflammatory and antioxidative component, resulted in reduced airway wall thickening in vivo and weakened cell viability, proliferation and migration capacity of ASMCs in vitro as well as a downregulation of the lncRNA PVT1.65 Short hairpins against PVT1 revealed its impact on cell viability, proliferation and migration capacity of ASMCs. Luciferase reporter assays showed that PVT1 acts as a sponge for microRNA-203a, which targets E2F3. Moreover, Yu et al demonstrated that this PVT1/miR-203/E2F3 pathway is affected by the α-asarone treatment. To conclude, results suggest that PVT1 is a sponge for microRNA-203a, which regulates the expression of E2F3 in these asthmatic rats.65 However, further research is needed to investigate if similar mechanisms occur in humans with asthma.
Wei et al investigated the function of PVT1 in mice with ozone-triggered asthma. They could link high PVT1 expression levels to asthma and especially thicker ASMCs. In in vitro experiments, PVT sponges microRNA-29c-3p and therefore induces PI3K–Akt–mToR signalling.66
Ma et al also investigated the function of PVT1 showing that overexpression led to pathological development in human small airway epithelial cells. PVT1 was also seen to sponge microRNA-149.67
Other lncRNAs
More lncRNAs have been studied and found to be upregulated or downregulated in asthma or a certain asthma type.11 12 38 68–78 A summary of these can be found in table 2, and a more extensive description of some of them can be found in supplemental materials. However, these lncRNAs were not extensively studied or described only in animals. As a result, more in vitro and in vivo experiments are needed to distinguish the exact function of these lncRNAs.
Conclusion
This review sheds light on lncRNA expression profiles in various cell types involved in the pathophysiology of asthma and their potential functional roles. Considering the heterogeneity of asthma and the published reports thus far, replication and validation studies on larger sample numbers from well-characterised patients are required to identify those lncRNAs that have a robust association with a specific asthma phenotype. The fact that differentially expressed lncRNAs can be detected in easily accessible samples such as plasma or serum underscores their potential to serve as biomarkers.11 12 38 45 Nevertheless, more studies are required to identify those lncRNAs profiles that can identify a specific phenotype or can predict therapeutic response.
At this stage, most publications are descriptive and do not include functional experiments, with only a handful of lncRNAs that have been functionally investigated, such as MEG3,34 LNC_0088246 and TUG1.57 There are thus multiple opportunities for further research.
Although lncRNAs can modulate gene expression in several ways (figure 1), most published functional studies in asthma focused on their capacity to sponge miRNAs. Further research is needed to investigate how the other lncRNA functionalities (decoys, guides and scaffolds) can have an impact on asthma and may present novel therapeutic opportunities. More multiomics approaches, combining genetics with lncRNA and miRNA expression data and proteomics, are needed to obtain insights into how genetics is linked to lncRNA and miRNA expression and how this impacts on protein expression in asthma. Little is known regarding the presence of lncRNAs in asthma genome-wide association study (GWAS) hits. To our knowledge, only one recent GWAS study reported an lncRNA (CTD-253719.5) that colocalises with a locus linked to lung function.79 Also, the working mechanisms of lncRNAs,for example, the protein interaction partners of lncRNAs, should be investigated with relevant technologies, such as ChIRP-MS (comprehensive identification of RNA-binding proteins by mass spectrometry, a technique to discover proteins that bind to an RNA of interest, figure 3).80
Still, several of the identified lncRNA-associated pathways have an impact on asthma-associated pathogenic processes, such as Th1/Th2 balance,38 47 48 51 airway remodelling41 46 50 57 65 74 76 78 and insensitivity against certain drugs,49 62 suggesting that lncRNAs could become therapeutic targets. However, translation of these insights into the clinic will require major technical breakthroughs, such as approaches to modulate lncRNA function in specific cell types within the human lung.
To conclude, lncRNAs hold great potential as both biomarkers and therapeutic targets in asthma. However, this potential remains far from being fully evaluated, and more research is needed to precisely determine, characterise and exploit the most promising candidates. Nevertheless, these preliminary studies argue in favour of systematically integrating lncRNAs in asthma research.
Ethics statements
Patient consent for publication
Ethics approval
This study does not involve human participants.
References
Footnotes
Twitter @Fien_G_
Contributors FG collected studies and wrote the review. EdBdL, PM and TM supervised literature review and writing. EdBdL and TM are joint last authors.
Funding The Department of Biomolecular Medicine is funded by a Concerted Research Action of Ghent University (BOF19-GOA-008). The Department of Respiratory Medicine is funded by a Concerted Research Action of Ghent University (BOF19-GOA-008) and Fund for Scientific Research in Flanders (FWO-EOS project G0G2318N).
Competing interests TM reports grants and personal fees from GlaxoSmithKline outside the submitted work, is a shareholder from Oryzon Genomics and Mendelion Lifesciences SL and holds a Chiesi chair on environmental factors in asthma.
Provenance and peer review Not commissioned; externally peer reviewed.